
Pelagic Zone
IV. Zooplankton
 Zooplankton are the major trophic link to pelagic vertebrates, and their
abundant populations draw many birds, fish and whales to the area. Whereas
phytoplankton production is limited to upper, sunlit waters, zooplankton
growth occurs at all depths, including the epipelagic (0-200 m), mesopelagic
(200-100 m) and bathypelagic (>1000 m) zones. In the MBNMS, zooplankton
are usually most abundant in neritic and inshore regions (Colebrook 1977),
as compared with waters of the offshore California Current. Biomass patterns
appear strongly linked to mesoscale circulation features. For instance,
the eddy/meander off Monterey Bay may be a focus for zooplankton (Huntley
et al. 1995), and such mesoscale features may show predictable gradients
in zooplankton species (Mackas et al. 1991). A strong seasonal pattern is
evident in zooplankton biomass, with highest amounts inshore and maxima
in spring and summer (U.S. GLOBEC 1994). The seasonal input of larvae, many
from shallow benthic invertebrates, is important regionally; it is likely
that spawning is triggered by phytoplankton blooms (Starr et al. 1990).
(Local return and settlement of larvae, however, may be strongly dependent
on mesoscale current features [Farrell et al. 1991, Graham 1994]). In additional
to the onshore-offshore, mesoscale, and seasonal variations, there is also
the well known decline in zooplankton abundance from the epipelagic to the
mesopelagic zone and below (Vinogradov and Tseitlin 1983). However, in the
epi- and mesopelagic zones these are complicated by daily vertical migrations
of many species. Vertical distributions of local plankton also are known
to be influenced by the intensity of turbulence (Haury et al. 1992)
Zooplankton are the major trophic link to pelagic vertebrates, and their
abundant populations draw many birds, fish and whales to the area. Whereas
phytoplankton production is limited to upper, sunlit waters, zooplankton
growth occurs at all depths, including the epipelagic (0-200 m), mesopelagic
(200-100 m) and bathypelagic (>1000 m) zones. In the MBNMS, zooplankton
are usually most abundant in neritic and inshore regions (Colebrook 1977),
as compared with waters of the offshore California Current. Biomass patterns
appear strongly linked to mesoscale circulation features. For instance,
the eddy/meander off Monterey Bay may be a focus for zooplankton (Huntley
et al. 1995), and such mesoscale features may show predictable gradients
in zooplankton species (Mackas et al. 1991). A strong seasonal pattern is
evident in zooplankton biomass, with highest amounts inshore and maxima
in spring and summer (U.S. GLOBEC 1994). The seasonal input of larvae, many
from shallow benthic invertebrates, is important regionally; it is likely
that spawning is triggered by phytoplankton blooms (Starr et al. 1990).
(Local return and settlement of larvae, however, may be strongly dependent
on mesoscale current features [Farrell et al. 1991, Graham 1994]). In additional
to the onshore-offshore, mesoscale, and seasonal variations, there is also
the well known decline in zooplankton abundance from the epipelagic to the
mesopelagic zone and below (Vinogradov and Tseitlin 1983). However, in the
epi- and mesopelagic zones these are complicated by daily vertical migrations
of many species. Vertical distributions of local plankton also are known
to be influenced by the intensity of turbulence (Haury et al. 1992)
A. Shallow Zooplankton
 The taxonomic composition of many zooplankton groups has been documented
regionally. Dominant epipelagic groups inshore are crustacean larvae, copepods,
euphausiids, ctenophores, hydrozoan medusae and siphonophores, and the chaetognaths
(Bigelow and Leslie 1930, Colebrook 1977). In most cases, the numerical
and biomass dominants are often copepods. These include small, neritic surface-dwelling
Acartia and Paracalanus, large, more oceanic Calanus pacificus,
vertically migrating Metridia pacifica and Pleuromamma and
deeper, nonmigrating Oithona (Haury et al. 1990, Huntly et al. 1995, Steinberg
unpublished data). Copepods appear to be separated to some extent into niches
that are onshore-offshore, depth and migration related, but patterns have
not been well described locally, though they likely resemble those described
to the north (Peterson et al. 1979).
The taxonomic composition of many zooplankton groups has been documented
regionally. Dominant epipelagic groups inshore are crustacean larvae, copepods,
euphausiids, ctenophores, hydrozoan medusae and siphonophores, and the chaetognaths
(Bigelow and Leslie 1930, Colebrook 1977). In most cases, the numerical
and biomass dominants are often copepods. These include small, neritic surface-dwelling
Acartia and Paracalanus, large, more oceanic Calanus pacificus,
vertically migrating Metridia pacifica and Pleuromamma and
deeper, nonmigrating Oithona (Haury et al. 1990, Huntly et al. 1995, Steinberg
unpublished data). Copepods appear to be separated to some extent into niches
that are onshore-offshore, depth and migration related, but patterns have
not been well described locally, though they likely resemble those described
to the north (Peterson et al. 1979).
Euphausiids (krill) are particularly important in food webs of the MBNMS and are major prey for marine vertebrates, including anchovy, squid, salmon, hake, blue sharks and many other vertebrates (Morejohn et al. 1978). Krill swarms, especially of the more oceanic Euphausia pacifica and the more neritic Thysanoessa spinifera draw a variety of predators. Dense aggregations can exist at depth, for example along the walls of Monterey submarine canyon, where they attract diving blue whales (Schoenherr 1991). Though krill are normally found at depth during the day, surface swarms occasionally occur that may be related to reproduction (Smith and Adams 1988). Surface patches, sometimes colored, are particularly important to seabirds and cetaceans (also see Seabird & Shorebird and Marine Mammal sections.) Distributions of krill and other zooplankton recently have been linked to mesoscale features (Huntley et al. 1995).
 "Jelly" species are conspicuous in MBNMS zooplankton communities,
especially since SCUBA divers, manned submersibles (Barham 1956) and MBARI's
remotely operated vehicle (ROV: Robison 1994)) have provided visual access
to the Bay's fauna. Surface water communities include a host of seasonally-produced
(mostly spring-summer) hydrozoan cnidarians, which are millimeter- to centimeter-sized
jellyfish that prey heavily on small zooplankton and are often budded from
benthic polyps (Smith 1977, Morris et al. 1980, Wrobel 1990). Other larger
scyphozoan medusae, with bells sometimes exceeding 10 or 20 cm in diameter,
occasionally swarm in the area. Abundant species, with tentacles and mouth
lobes trailing sometimes meters below the swim bell, include the purple
striped jelly Pelagia, the sea nettle Chrysaora, and the abundant
moon jelly, Aurelia, all important predators on copepods, fish larvae
and other zooplankton (Morris et al. 1980; Wrobel 1990). The first two of
these occasionally wash up on local beaches. When abundant in summer, these
scyphozoans may draw sea turtles to this region (Starbird et al. 1993).
The large jellies often are colonized by juvenile crustaceans, including
the megalopae of Cancer crabs. They are sometimes held offshore during upwelling
by fronts (Wrobel 1990; Graham 1994).
"Jelly" species are conspicuous in MBNMS zooplankton communities,
especially since SCUBA divers, manned submersibles (Barham 1956) and MBARI's
remotely operated vehicle (ROV: Robison 1994)) have provided visual access
to the Bay's fauna. Surface water communities include a host of seasonally-produced
(mostly spring-summer) hydrozoan cnidarians, which are millimeter- to centimeter-sized
jellyfish that prey heavily on small zooplankton and are often budded from
benthic polyps (Smith 1977, Morris et al. 1980, Wrobel 1990). Other larger
scyphozoan medusae, with bells sometimes exceeding 10 or 20 cm in diameter,
occasionally swarm in the area. Abundant species, with tentacles and mouth
lobes trailing sometimes meters below the swim bell, include the purple
striped jelly Pelagia, the sea nettle Chrysaora, and the abundant
moon jelly, Aurelia, all important predators on copepods, fish larvae
and other zooplankton (Morris et al. 1980; Wrobel 1990). The first two of
these occasionally wash up on local beaches. When abundant in summer, these
scyphozoans may draw sea turtles to this region (Starbird et al. 1993).
The large jellies often are colonized by juvenile crustaceans, including
the megalopae of Cancer crabs. They are sometimes held offshore during upwelling
by fronts (Wrobel 1990; Graham 1994).
Other jelly predators include the siphonophores. These are colonial hydrozoan jellyfish that range from the surface to the deepest waters of the MBNMS. Important surface forms include Muggiaea and other species (Bigelow and Leslie 1930), but the group is poorly known from conventional plankton net samples because the delicate colonies often are damaged beyond recognition during capture. Surface species often have small gas-filled floats and prey on zooplankton.
The comb jellies, members of the phylum Ctenophora, include important local species, mostly poorly known because of their fragility (Wrobel 1990). The sturdiest of these is Pleurobrachia. An occasional dominant in the spring and summer, it consumes a variety of small prey, especially copepods (Bigelow and Leslie 1930). Pleurobrachia can occasionally be found in strand lines along the area beaches, looking like centimeter-sized, colorless marbles. Others comb jellies, like Leucothea, are extraordinarily fragile (Wrobel 1990) and reported mostly by divers.
A particularly abundant jelly in the region is the By-the-wind-sailor, Velella velella, a sail-bearing cnidarian that blows ashore in windy periods in spring and summer (Francis 1985, Wrobel 1990). The lovely, small blue floats possess dinoflagellate (algal) symbionts, but also consume small zooplankton. The bleached floats can persist in drifts for months (also see Sandy Beach section).
 Locally, larvaceans (gelatinous invertebrate chordates) can be important
in the epipelagic zone (Bigelow and Leslie 1930). These organisms are generally
small (millimeter-sized) and tadpole-like in appearance; they inhabit transparent
feeding devices, called "houses" which are food-concentrating
structures (Alldredge 1976). Several species can be abundant locally, and
one - Oikopleura longicauda - occasionally occurs in long,
red, surface windrows (Alldredge 1982, M. Silver unpublished data). Larvaceans
discard their millimeter- to centimeter-sized houses every few hours (Alldredge
1972), contributing importantly to "marine snow", the visible,
non-living particles found abundantly in the MBNMS (Silver et al. 1978).
Along with phytoplankton, marine snow reduces visibility for recreational
divers during much of the year, especially during the upwelling season.
Locally, larvaceans (gelatinous invertebrate chordates) can be important
in the epipelagic zone (Bigelow and Leslie 1930). These organisms are generally
small (millimeter-sized) and tadpole-like in appearance; they inhabit transparent
feeding devices, called "houses" which are food-concentrating
structures (Alldredge 1976). Several species can be abundant locally, and
one - Oikopleura longicauda - occasionally occurs in long,
red, surface windrows (Alldredge 1982, M. Silver unpublished data). Larvaceans
discard their millimeter- to centimeter-sized houses every few hours (Alldredge
1972), contributing importantly to "marine snow", the visible,
non-living particles found abundantly in the MBNMS (Silver et al. 1978).
Along with phytoplankton, marine snow reduces visibility for recreational
divers during much of the year, especially during the upwelling season.
Other species of gelatinous plankton visit surface waters of the MBNMS, often in association with inshore movements of more oceanic, California Current waters. These visitors include salps and doliolids, pteropods (sea butterflies) including the larger mucus web feeders (e.g. Corolla) as well as the smaller carnivores (e.g. Clione) (regional offshore distributions in CalCOFI Atlases, condensed in Colebrook 1977, unpublished inshore obs. by authors).
Changes in the local currents, especially those associated with ENSO (El Niño-Southern Oscillation) events, can dramatically alter both the abundance and species composition of the zooplankton in upper waters. Thus, the presence of the swimming red crab Pleuroncodes is a sure sign of southern water, as this species normally lives off Baja California (McGowan 1984). Likewise, the decline of temperate species including local krill, a change that occurs with warming events like ENSO, may negatively effect marine birds and mammals. Decadal scale changes also have been reported in zooplankton abundance in the in southern California (Roemmich and McGowan 1995), but such changes are not as so clear in the MBNMS area (Chelton et al. 1982). Decadal scale changes have been linked less with alterations in upwelling intensity than with variations in the water flow rates of the southward-moving California Current (Chelton et al. 1982, Roemmich and McGowan 1995).
B. Deep Zooplankton
 At depth, the zooplankton fauna is much less well known. Observations
from ROVs (Remotely Operated Vehicles) and submersibles have been particularly
important in the MBNMS in discovering new species, learning more about previously
named species, and studying behavior. One of the more common deepwater hydrozoans
(a narcomedusa) is Solmissus , a predator on other gelatinous zooplankton
(Mills and Goy 1988). The siphonophores are particularly prominent in deep
waters and are important predators. Some, such as Apolemia, may
reach 40 meters in length in the Monterey Submarine Canyon (Barham 1956,
Robison 1995).The fragile, deep water ctenophore fauna was virtually unknown
until the studies using MBARI's ROV. Matsumoto and Robison (1992) recently
described Kiyohime usagi, the large (up to 28 cm) , "rabbit-eared"
ctenophore. It is a large, passive predator that ambushes euphausiids. Other
poorly known ctenophores of the MBNMS's mesopelagic and bathypelagic waters
have also been recorded recently by Matsumoto (1990).
At depth, the zooplankton fauna is much less well known. Observations
from ROVs (Remotely Operated Vehicles) and submersibles have been particularly
important in the MBNMS in discovering new species, learning more about previously
named species, and studying behavior. One of the more common deepwater hydrozoans
(a narcomedusa) is Solmissus , a predator on other gelatinous zooplankton
(Mills and Goy 1988). The siphonophores are particularly prominent in deep
waters and are important predators. Some, such as Apolemia, may
reach 40 meters in length in the Monterey Submarine Canyon (Barham 1956,
Robison 1995).The fragile, deep water ctenophore fauna was virtually unknown
until the studies using MBARI's ROV. Matsumoto and Robison (1992) recently
described Kiyohime usagi, the large (up to 28 cm) , "rabbit-eared"
ctenophore. It is a large, passive predator that ambushes euphausiids. Other
poorly known ctenophores of the MBNMS's mesopelagic and bathypelagic waters
have also been recorded recently by Matsumoto (1990).
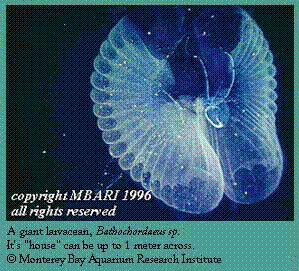
A number of species are now being studied extensively using the MBARI ROV. One example is the yellow mesopelagic worm Poeobius, a mesopelagic detrital feeder that uses mucus threads to capture sinking detritus (Utall and Buck 1996) Another example is the mesopelagic "giant larvacean" Bathochordaeus that builds houses up to 1 m across (Hamner and Robison 1972); these houses support a unique fauna of associated zooplankton (Steinberg et al. 1994). Bathochordaeus occurs in the high salinity California Undercurrent and is known from more southerly waters (Barham 1979). After being discarded, the large houses sink rapidly, carrying shallower materials to the seafloor below (Hamner and Robison 1992).
 Below 500 m, most plankton are colored deep reds, browns, purples and black:
these color changes are generally associated with the lack of light, especially
red light, at depth. (Illuminated by the faint blue-greenish light that
penetrates deepest, red and dark colored objects are non-reflective and
invisible.) This is the zone whose fauna is poorly known, because of fragility
of individuals (resulting in unrecognizable specimens from net tows) and
low abundance. Forms include the archaic, football-sized "vampire"
squid Vampyroteuthis infernalis, and the beautiful red/purple medusa
Atolla (Robison 1995). Vampyrocrossota childressi, the only
known black medusa, occurs in the Monterey Canyon at depths from 600-1500
m (Thuesen 1993). Many of the mesopelagic and bathypelagic species are bioluminescent,
using body lights and extruded glowing clouds for various purposes (Robison
1995).
Below 500 m, most plankton are colored deep reds, browns, purples and black:
these color changes are generally associated with the lack of light, especially
red light, at depth. (Illuminated by the faint blue-greenish light that
penetrates deepest, red and dark colored objects are non-reflective and
invisible.) This is the zone whose fauna is poorly known, because of fragility
of individuals (resulting in unrecognizable specimens from net tows) and
low abundance. Forms include the archaic, football-sized "vampire"
squid Vampyroteuthis infernalis, and the beautiful red/purple medusa
Atolla (Robison 1995). Vampyrocrossota childressi, the only
known black medusa, occurs in the Monterey Canyon at depths from 600-1500
m (Thuesen 1993). Many of the mesopelagic and bathypelagic species are bioluminescent,
using body lights and extruded glowing clouds for various purposes (Robison
1995).
| < Previous Section III. Phytoplankton |
||